Please note that this material is copyright to the publishers of the SPUMS Journal and to the author
Not to be reproduced in any way without permission except for personal study
SPUMS J, 23(2), 1993, 93-102
Production of geological structures by the green alga Halimeda
Edward A. Drew
Australian Institute of Marine Science. PMB No 3, MC, Townsville, Q 4810. Australia.
Halimeda is a genus of calcareous geeen algae found throughout the tropics, mainly on coral reefs. One species also occurs in the subtropics and another in the Mediterranean. Twenty of the world's 30 Halimeda species grow, often prolifically, on the Great Barrier Reef(GBR). Most of those not found there are confined to the Caribbean, having evolved there after the closure of the Isthmus of Panama in the Miocene, 20 million years ago.
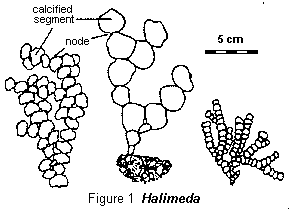
The appearance of Halimeda plants is shown in Figure 1. They are all composed of numerous flat segments between 0.5 and 3 cm wide, depending on species. These segments are calcified, sometimes very heavily, and they are joined by very short, uncalcified nodes to form branching plants. Studies of these algae have, until recently, concentrated on the prodigious amounts of coarse calcareous sediments they produce on coral reefs when they die and then quickly disintegrate at the nodes to produce piles of calcified segments. However, during the last 10 years, algologists and geologists have combined to show that Halimeda can grow and produce sediment even more prolifically on the seabed away from reefs. This work began in the GBR but similar phenomena are now being studied both in Indonesian waters and as far away as the Caribbean.
There are more than 2,000 individual coral reefs scattered throughout the 268,000 km2 covered by the GBR. The reefs themselves cover only about 13,000 km2 leaving a lot of non-reefal seabed in between. This seabed slopes gently from the shoreline to depths of 50, or occasionally 100 m, at the outer edge which can 100 km or more offshore. The outermost reefs occur at the very edge of the continental shelf where the slope of the seabed suddenly increases dramatically and rapidly descends to 1,000 m.
The coral reefs of the GBR have been studied much more than the inter-reefal water mass and seabed. Hardly any attention was paid to this enormous area until marine scientists began to suspect that individual reefs did not behave as independent entities. Intensive study of the Crown of Thorns starfish infestations, which have plagued the GBR for decades, has served to emphasise the interconnectedness of reefs over long distances. What happens in the inter-reefal water connecting the reefs has now assumed vital importance.
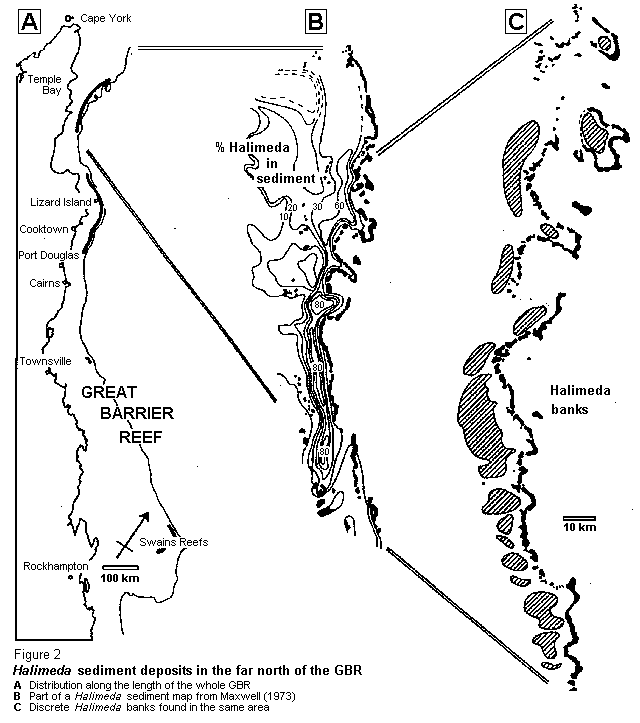
One researcher in particular provided fundamental information about the inter-reefal seabed well before this part of the GBR hecame a focus of scientific attention. Over 25 years ago Graham Maxwell, a geologist, organised a series of research cruises to sample and characterise the seabed sediments between the reefs throughout the region. This involved more than 6,000 grab sampling stations and a prodigious amount of sediment sieving and particle analysis. His work initially concentrated on the southern half of the GBR and the results were included in his Atlas of the Great Barrier Reef (1). However, he extended his studies northwards and, in 1973, published a thorough description of the sediments of the inter-reefal seabed of the whole GBR (2). It was his maps showing large areas of Halimeda-dominated coarse gravels, particularly in the northern part of the GBR (Figure 2B), which first diverted our attention away from Halimeda on the reefs and into this much more intriguing environment.
From about Port Douglas north, the outer edge of the GBR consists of a continuous strip of coral reef dissected every few kilometres by narrow passages about I km wide. The resulting string of long, narrow ribbon reefs forms an effective barrier between the Coral Sea and the waters on the continental shelf. This barrier extends for nearly 1.000 kilometres. Maxwell's studies. since supplemented by others (3,4), revealed that Halimeda gravels form major sediment deposits in a strip a few kilometres wide just behind this outer barrier and along most of its length. Our recent discovery of similar sediment behind Escape and Agincourt Reefs, just north of Port Douglas, extends the known Halimeda-rich deposits to the very bottom of the ribbon reefs.
The rest of the inter-reefal sediments of the northern GBR are, in the main, muddy and contain varying amounts of debris from the hard body parts of such organisms as molluscs and echinoderms. However. within a few kilometres of the reefs themselves the sediments turn into sand which consists mainly of coral fragments. It seems that such reefal debris is seldom transported very far from the reefs. Halimeda is typically a plant of shallow water on coral reefs. It was, therefore, surprising to find such large amounts of Halimeda debris several kilometres from the nearest reefs. particularly as those deposits are always separated from the nearby reefs by expanses of either coral sand or mud.
Orme et al (3) hinted at an explanation when they mentioned, almost casually, seeing large areas of luxuriant Halimeda plants on the deposits near Lizard Island. They probably also saw at least as much Halimeda sediment devoid of vegetation as they saw covered by algae and, as geologists, they did not pursue this botanical observation any further.
In 1983 the opportunity arose for us to work from HMAS KIMBLA, a boom defence vessel used by the Navy for hydrographic work. During that cruise we were able to visit not only the Lizard Island Halimeda gravel deposits but also two other areas further north. Despite atrocious weather with 40 knot winds, we were able to sample the seabed with grab and dredge at all three localities, and even managed to dive every day to photograph and sample directly the luxuriant Halimeda vegetation. Living Halimeda did indeed cover wide areas of the seabed and it was growing on sediment consisting of up to 96% Halimeda fragments. The unprecedented success of that, my first geo-botanical research cruise, is almost entirely due to the massive proportions and draught of HMAS KIMBLA which travelled sedately through very rough seas at about 6 knots and hove to for sampling with virtually no motion!
Quite clearly, the extensive deposits of Halimeda gravel, which seem to interest sedimentary geologists just as much as algologists were being generated in situ by luxuriant meadows of living algae. We have now studied the outer GBR in considerable detail from the northern limit of the GBR Marine Park, level with Cape York, to the bottom of the ribbon reefs off Port Douglas (5,6). These surveys have confirmed Maxwell's map (Figure 2C), with the notable exception of the large area he showed in the far north. Maxwell took only a few samples there and he was not to know that this area, which has the usual barrier of ribbon reefs on the map, has a sufficiently different hydrographic structure to preclude the formation of Halimeda meadows and gravel deposits.
The extent of the Halimeda meadows in the GBR is indicated in Figure 2A. They mostly reach to within 30 m of the surface and they essentially cease at Agincourt Reet. There are a few isolated meadows in the Townsville region which extend to at least 95 m in deep water near the shelf break and we have also investigated a few on the top of shallow reefal platforms in the Swains reefs at the southern end of the GBR. Neither of those rather different situations will be discussed here.
One important lesson we learned early during these surveys was that Halimeda meadows are extremely difficult to locate by grab sampling but they are extraordinarily easy to locate using the ship's echo-sounder even when steaming at 8 to 10 knots. This is because they are not, as our initial dives had indicated, flat expanses of wall-to-wall algae. They are actually composed of many small mounds, just a few hundred metres in diameter and up to 20 m high (Figure 3B), although these may sometimes grow together to form ridges.
This vertical relief still puzzles us somewhat but there are distinct clues as to the origin of the mounds and what keeps them as such. There is usually little or no living Halimeda on the sediment in the hollows between the mounds, so the meadows are seldom continuous from one mound to the next. This has been confirmed by observations from the Australian-built Platypus submersible (7) off Cooktown and also by underwater video transects we have surveyed throughout the GBR. The submersible cruises also found pinnacles of old, eroded coral rock between the mounds. These had some living corals on them but the rock was found to be of Pleistocene age. They must, therefore, have been exposed to aerial weathering during the last ice-age when sea-level was as much as 80 m lower than now and the GBR was dry land. We have also encountered similar lumps of coral rock in the depressions between mounds on many of our video-transects. In fact, they are a major hazard to the towed video camera which only survives because it is inside a heavy duty mesh cage!
The mounds probably originated as isolated patches of sediment between the pinnacles as sea level began to rise after the ice-age 10,000 years ago. Those patches would have had little Halimeda on them initially but the algae would have begun to grow as the water got deeper. Herbivorous fish ranging out from the coral pinnacles may then have kept nearby sediment free of Halimeda vegetation. Prolific sediment production by the ungrazed patches of vegetation further from the pinnacles would then, over thousands years, produce mounds capped by Halimeda meadow.
Whatever the reasons for this vertical irregularity of the Halimeda gravel deposits, there is no doubt such deposits cover considerable areas of seabed. In the northern GBR they may cover up to 2,000 km2, a substantial area approaching half that of the reefs themselves. We have been able to map these accumulations in detail because of their distinctive echo-sounder signature. They form discrete banks, several kilometres long, behind each ribbon reef, with a more or less distinct break associated with each major passage through the outer barrier (Figure 3A). This has led us to subdivide the larger areas of Halimeda gravel into Halimeda banks, which can be conveniently named after the ribbon reef they lie behind. These banks are composed of numerous mounds of gravel themselves covered by dense meadows of sediment-generating Halimeda plants. However, this is far from the end of the story, for this unique ecosystem has much more to tell us.
The teams led by Orme, working near Lizard Island, and by Davies, working off Cooktown, both carried out extensive sub-bottom seismic profiling. They showed that Halimeda sediment is extremely uniform in seismic reflectance and, although it was not very dense as compared with nearby coral sands, a distinct layered structure was clearly visible. These features are illustrated in Figure 3C, which also shows that the Halimeda gravels form a layer up to 20 m thick on top of the Pleistocene discontinuity, a particularly reflective structure which was formed during aerial exposure during the last ice age.
Cores taken by the geologists through the upper 5 m of the Halimeda sediment have confirmed their uniform, Halimeda dominated, composition although no structure has been detected which would explain the layered appearance on seismic profiles. Because 5 m is the deepest core which can he taken with current vibrocoring equipment and the deposits are 15 to 20 metres thick, it was necessary to core on the edges of the deposits and in the depressions between the mounds in order to sample the bottom layers. Such cores indicate that the older sediments near the bottom contained less Halimeda and more fragments of other calcareous organisms, especially the skeletons of the disc-shaped calcareous protozoa called foramenifera. Even more significantly, these sediments rest directly on a thin layer of mangrove peat, deposited when this part of the GBR seabed was the continental shoreline!
Carbon-14 dating of the Halimeda fragments at the bottom of 5 m cores shows them to be 3,000 to 5,000 years old, indicating a vertical accretion rate of up to 1m per thousand years.
Fortunately, we had already investigated the rate at which Halimeda vegetation dies, disintegrates and turns to sediment on reefs. By following the growth and loss of tagged parts of plants (8) we found that quite modest Halimeda vegetation, with 1 kg of plants per m2 could generate at least 2 kg of calcareous sediment per year. The species composition and rates of photosynthesis determined for the inter-reefal meadows was very similar to those found on the reefs, so we can confidently extrapolate the reef results to the inter-reefal situation. As the density of the Halimeda gravels from vibrocores was abut 0.7 g per cm3, one kg of calcareous debris spread over a square metre would raise the seabed by about 2.8 mm per annum, more than enough to account for the thickness of sediment now present. Some areas of Halieda vegetation actually have more than 3 kg of plants per m2, and so could generate sediment even more rapidly. There can, therefore, be little doubt that in-situ meadows of Halimeda can generate large sediment masses unaided. Indeed, they clearly rival the reefs themselves in laying down massive calcium carbonate structures for inclusion in the geological record.
Why do Halimeda banks grow only in the lee of the ribbon reefs
We have ascertained that the relatively insubstantial alga Halimeda could and almost certainly has generated these large structures. We can see the meadows and sediment banks on one series of standard aerial photographs of the GBR taken when the water was particularly clear. The banks have even been assigned numbers in the same sequence as the real coral reefs on the Great Barrier Reef Marine Park Authority zoning maps! Those photographs were initially mis-interpreted as an eroded karst topography (9). Hopley saw the dark circular patches as erosion hollows, presumed to have been generated when the shelf was last exposed, but our echo-sounding surveys and diving confirm that the dark areas are Halimeda meadows atop mounds of sediment and the light areas are either the unvegetated hollows between them or the tops of currently unvegetated mounds. It is even possible, as will be explained below, that we can see the chlorophyll within the plants on images from satellites in space.
We must now ask why Halimeda banks occur only in that narrow belt a few kilometres behind the outer barrier reefs. More directly, we might ask how can a luxuriant algae vegetation develop and thrive for thousands of years beneath waters virtually devoid of essential nutrients. Algae, like most plants, require both inorganic nitrogen (nitrate, nitrite or ammonnium ions) with which to synthesis new protein for growth, and they also need phosphate to support their complex biochemistry. The levels of these nutrients in the shallow shelf waters (0.04 mM nitrate,0.07 mM ammonium and 0.14 mM phosphate) are insufficient to support any significant algal growth. However we also know there is more than enough of these nutrients tantalisingly close, for in the adjacent Coral Sea we find 0.7mM phosphate and nitrate levels as high as 8 mM, but only at depths greater than 8O to lOOm.
The reason these nutrients are in short supply in surface waters even in the Coral Sea is that biological productivity binds them into living organisms. These then die and sink below the depths reached by the mixing processes driven by wind and tide. This leaves the upper, mixed layer of the ocean depleted of nutrients and therefore relatively unproductive. Because the passages between the reefs are typically about 45 m deep. tidal exchange cannot be expected to transport water other than that from the mixed layer onto the shelf, so it is not surprising the shelf waters are also nutrient depleted.
But things are not quite w hat they seem on the surface. Firstly. there are the strong tidal currents in the passages through the outer barrier to connsider. Research elsewhere on the GBR had shown that the deeper water in these passages is somewhat colder than the rest of the mixed layer. Oceanographers generally associate cold water with nutrient-rich water from beneath the thermocline, which is also situated at 80 to 100m depth in the Coral Sea. Perhaps we do, after all, have a potential source of nutrients for bottom-dwelling algae.
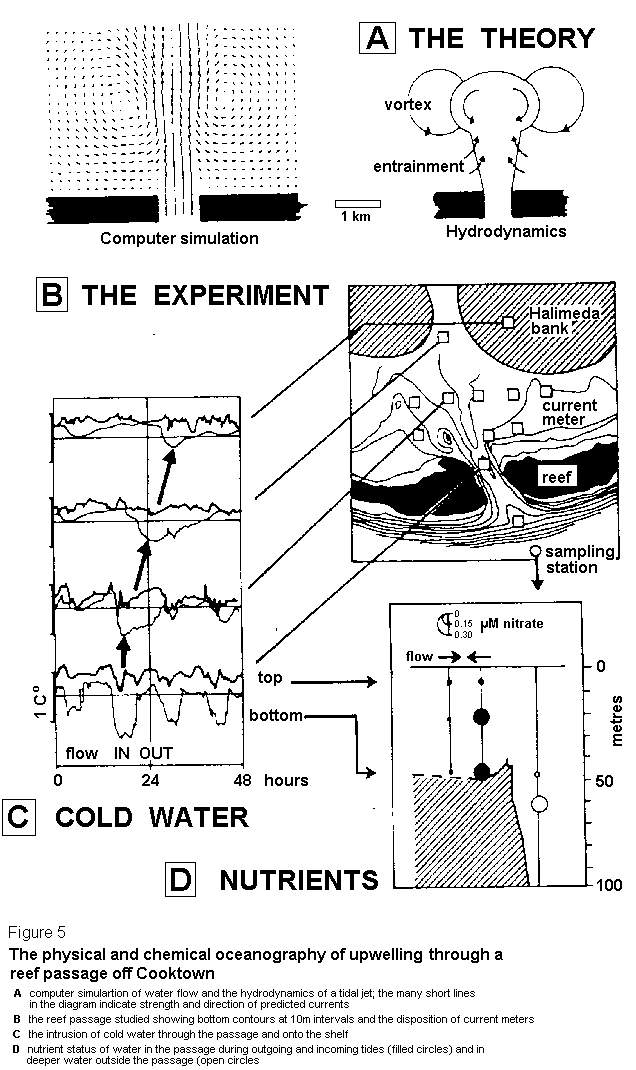
The second part of this equation comes from classical hydrodynamics which predict that a strong flow through a narrow opening will continue as a discrete jet far beyond that opening. Computer simulations of the reef passage situation confirmed this possibility and also indicated that a few kilometres inside the opening the flow should slow and separate into two rotating vortices situated precisely where the Halimeda banks grow (Figure 5A).
We carried out a large multidisciplinary experiment to test the hypothesis that strong tidal currents caused nutrient upwelling through the reef passages. This experiment involved a dozen current meters, a CTD profiler able to measure salinity and temperature to several hundred metres depth, Nislkin bottles to collect water samples at similar depths, some surface drogues to follow water movement, and aerial photography of the jets from a light aircraft. That experiment (Figure 5B) showed that our computer predictions were indeed correct (10). On the incoming tide we detected cold water brought up from below the thermocline and propagated through the reef passage onto the shelf in a layer nearly 20 m thick (Figure 5C). Once through the passage, that water slowed down, formed vortices and eventually reached the Halimeda hanks 12 hours later. Considerable nitrate and phosphate enrichment of the water at the bottom of the passage was also detected (Figure 5D) but, unfortunately, we could not detect propagation of these nutrients far onto the shelf. Perhaps there was too much dilution with depleted shelf water or some of the nutrients were taken up by phytoplankton during their 12 hour journey from the shelf-break to the meadows. In any event, it was reassuring to observe that neither cold water nor nutrients were exported through the passage on the outgoing tide because, as predicted, the out-going water came from directly behind the ribbon reefs which had not been enriched at all by the tidal jet.
This mechanism effectively pumps a considerable amount of nutrient-rich sub-thermocline water onto the shelf every time the tidal currents are strong enough to lift water from below the thermocline, at about 8O m, over the 45 m deep sill of the reef passage.
ln the passage we studied. the tidal currents appear to be strong enough to do this on both tides each day for up to 3 days either side of high spring tides. We have evidence, albeit less complete. of this phenomenon in several other passages, so it probably occurs along the entire length of the ribbon reefs. However. the upwelling will only occur if the reef passage is at least 40 m deep. Most passages through the outer barrier are deep enough, but a few are not and passages less than 40 m deep to not have Halimeda banks associated with them. This further supports our hypothesis, and also explains the absence of Halimeda gravel deposits in the extreme north where Maxwell predicted them but we could find none, for here none of the reef passages are more than 30 m deep.
We have concentrated here on events during the incoming tide. A similar tidal jet and upwelling from below the thermocline also occurs in the Coral Sea during the outgoing tide. This process effectively enriches the surface waters just outside the ribbon reef and may directly benefit those reefs. It may also not be coincidence that boats fishing for black marlin patrol exactly these outgoing tidal jets and associated vortices.
Our calculations of inorganic nitrogen fluxes indicate that 58 metric tonnes of nitrate are imported each year through a typical reef passage 40 m deep and 1 km wide. The 15 km2 of Halimeda meadow associated with each such passage actually required about 48 metric tonnes per year so such upwelling would be sufficient to allow algae to thrive behind the ribbon reefs below shelf waters otherwise too depleted of nutrients to support their growth. The reason the alga which grows there is usually Halimeda, and therefore can generate substantial sedimentary structures, is not so clear, whilst the absence of detectable nutrient enrichment over Halimeda banks themselves suggests that the processes involved may be less direct than we initially hypothesised.
We are now in the process of refining our hypothesis. Satellite images of the northern GBR indicate dramatic and dynamic accumulations of chlorophyll along both sides of the ribbon reefs (Figure 6A). As satellite cameras can certainly "see" 20 m or more through clear waters, some of this chlorophyll may actually be that in the benthic Halimeda vegetation. A single pixel of especially high chlorophyll almost exactly over the top of the shallowest mound we know, which is only 16 m below the surface, supports this possibility (see Figure 6C). Nevertheless, most of the chlorophyll undoubtedly represents phytoplankton growing in response to shelf-break upwelling events such as the one we have descrihed, and therein lies a possible solution to our dilemma.
Continuous recording of chlorophyll in surface waters has revealed that this can be transported from the Coral Sea onto the shelf through the reef passages, just as cold water and some nutrients are. Detailed examination of one such area showed plumes of cool water rich in particulate chlorophyll, i.e. phytoplankton, flowing through the passages and even splitting into a double peak suggestive of the twin vortices of a tidal jet (Figure 6D). Uptake by this phytoplankton could certainly account for the disappearance of some of the nutrients freshly upwelled into the reef passage before they could progress far onto the shelf. This phytoplankton, just like our cooler water and nutrients, passed in through the passages but not out again. So it could reside a few kilometres behind the reefs for some time, again just as our cooler water did. During that time the processes of zooplankton grazing and defaecation would certainly cause particulate, nutrient-rich material to fall on the Halimeda banks. awaiting only the final bacteria-mediated remineralisation before becoming available to the alga on the sea bed as inorganic nutrients.
Thus, we should perhaps add passage through phvtoplankton, grazing zooplankion and bacteria to the route we originally proposed for nutrients travelling from below the thermocline to the Halimeda banks. These processes await quantification but do promise even greater quantities of nutrients to support the banks than did our original model which took account only of the dissolved inorganic nitrogen upwelled through the passages. It now appears that nitrogen bound organically within the phytoplankton cells accompanies those nutrients and this has now been tracked all the way to the Halimeda baiks using the spectral signature of phytoplankton chlorophyll.
The biological, oceanographic and geological processes we have discovered whilst studying the Halimeda banks are helping us understand just how the Great Barrier Reef functions as an entity. However, they have a greater significance which extends beyond our region, for very similar deposits have been accumulating elsewhere for not thousands of years but hundreds of millions of years.
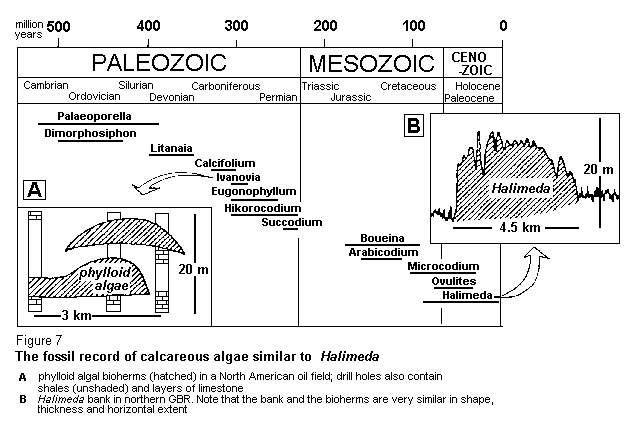
Halimeda is the most recent of a long line of foliose calcareous algae collectively recognised as phylloid algae in the fossil record (Figure 7). The internal organisation of the segments, which is so important for identifying living species of Halimeda, is mirrored in the calcium carbonate skeleton which becomes preserved intact in the sediment. Similar structures can be recognised in rocks hundreds of millions of years old, rocks which have been formed by lithification ofsediment deposits. These rocks tend to form lens-shaped domes several kilometres in extent and many metres thick, usually in the outer regions of ancient reefal systems. These reefal systems may pre-date the evolutionof the corals, but the phylloid algal bioherms they contain closely resemble the sediment deposits we have been studying in the northern GBR (Figure 7 A,B).
Why do we know so much about the ancient phylloid algal bioherms, and why have geologists laboured long and hard to understand just how and why they were formed? These deposits are porous, just like Halimeda gravel, because of all the small spaces retained within the skeletal fragments. And because of this porosity, they have come to form major oil reservoirs in many parts of the world. However, the Halimeda deposits of the Great Barrier Reef will not attract exploration for some time. The GBR is so young geologically that, even if its Halimeda banks do have the composition, texture and appearance of potential bioherms, none of them will become commercially interesting for millions of years.
Atlas of the Great Barrier Reef. Elsevier, Amsterdam.
2. Maxwell (1973)
Sediments of the Great Barrier Reef Province. In Biology and Ecology of Coral Reefs. Volume 1. Geology, edited Jones and Endean. Academic Press, New York and London: 299-345.
3. Orme, Flood and Sargent (1978)
Sedimentation trends in the lee of outer (Ribbon) reefs, northern region of the Great Barrier Reef. Phil Trans Roy Soc London, Series A; 291: 85-99.
4. Davies and Marshall (1985)
Halimeda bioherms - low energy reefs, northern Great Barrier Reef. Proc 5th Intl Coral Reef Congress, Tahiti. 1985; 5:1-7.
5. Drew and Abel (1985)
Biology, sedimentology and geography of the vast inter-reefal Halimeda meadows within the Great Barrier Reef Province. Proc 5th Intl Coral Reef Congress, Tahiti. 1985; 5:15-20.
6. Drew and Abel (1988)
Studies on Halimeda. l. The distribution and species composition of Halimeda meadows throughout the Great Barrier Reef. Coral Reefs; 6: 195-205.
7. Phipps, Davies and Hopley (1985)
The morphology of Halimeda banks behind the Great Barrier Reef east of Cooktown, Queensland. Proc 5th Intl Coral Reef Congress, Tahiti. 5: 27-30.
8. Drew(1983)
Halimeda biomass, growth and sediment generation on reefs in the central Great Barrier Reef Province. Coral Reefs. 2: 101-110.
9. Hopley (1978)
Geomorphology of the reefs and reef island, Great Barrier Reef north of Lizard Island. In Workshop on the northern section of the Great Barrier Reef GBRMPA, Townsville,219-252.
10. Wolanski, Drew, Abel and O'Brien (1988)
Tidal jets, nutrient upwelling, and their influence on the productivity of the alga Halimeda in the Ribbon reefs, Great Barrier Reef. Estuarine, Coastal & Shelf Sci. 26: 169-201.