Helga's retina
book:
Ron Douglas:
Contents and links:
Helga's retina
book:
Ron Douglas:
Contents and links:
Helga's retina
book:
Ron Douglas:
In order to get a more complete picture of the changing morphology within the same class of cell, we drew every single cell within several sections. A summary drawing is shown below.
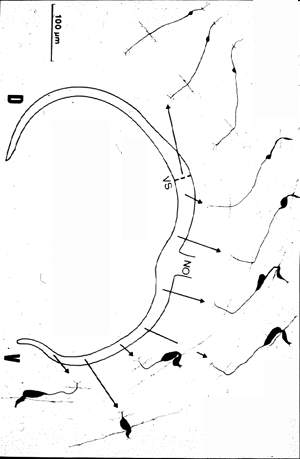 |
Summary drawing of the variability encountered
within the same class of retinal biplar cell,
depending on the retinal location. In the central
retina, the cells are numerous and small, whereas
in the periphery, the density decreases by a factor
of about 100, concomittant with an enormous
increase in size of the cell and the dendritic field.
Interestingly, the size of the output location does
not vary with retinal eccentricity
(VS: visual streak; D: dorsal; V: ventral) |
The first approach was to try staining a retina as a whole (retinal
wholemount). Unfortunately, this approach, although it resulted in extremely
beautiful views of the retinal surface, did not yield the anticipated data.
The photoreceptors themselves utilize colored oil droplets as color filters
to enhance color vision and these very oil droplets turned out to be a
major obstacle to the attempt of viewing the detailed pattern of stained
bipolar cell dendritic processes. Thus, the desired structures could, in
the best case only partially be detected
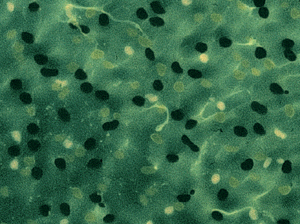
| Wholemount view of the turtle retina stained with an antibody against serotonin. Some labeled processes are faintly visible between the colored oil droplets of the photoreceptors. Note that because of the fluorescent illumination, the colors of oil droplets are not represented correctly, that is, in reality, the black droplets are red |
Another, quite a bit more difficult approach was to take the retina, flatten it and cut it tangentially, not an easy task, considering the fact that the entire layer of neural tissue is only 150 200 µm thick. Nevertheless, it is possible and the following pictures show a series of such sections through the different retinal layers.
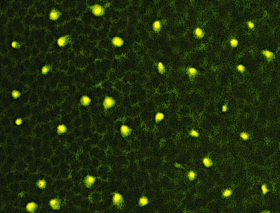
| Section through the outer nuclear layer, showing the honeycomb pattern of photoreceptors with labeled Landolt's clubs interspersed between them. |
Sections through the outer nuclear layer, show the mosaic of the photoreceptor
cells, visible as a honeycomb pattern of black circles. In between,
the bright yellow dots are sectioned Landolts clubs of bipolar cells stained
by the immunoreaction against serotonin. Sections like the one shown allowed
us to perform a quantitative analysis of the number of photoreceptors in
each given retinal area as well as a correlation of the number of stained
bipolar cells with the number of photoreceptors. Today, it is almost generally
accepted that this number is constant through the entire retina but 15
years ago, when this study was conducted, this finding was quite exciting.
The differential distribution of photoreceptors, depending on the retinal
topography had been known for almost 100 years but almost no further
data regarding other classes of retinal cell were available at the time,
for the simple fact, that no specific markers were available yet.
In the specific case of the turtle, which has about 100 times more
photoreceptors in the visual streak area than in the retinal periphery,
this meant that our preliminary assumption about bipolar cells showing
a similar density distribution was correct. It is well established that
humans as well as other species have a much lower visual acuity in the
retinal periphery. If this were not only caused by a lower number of photoreceptors
but also reflected in the numbers of all other retinal cells, including
bipolar and ganglion cells, one would only have to assume that each ganglion
cell occupies the same amount of territory in the brain. In other words,
only a few bipolar and ganglion per given area in the periphery as opposed
to a 100-fold higher number in the central retinal area would almost suffice
to provide the anatomic correlate to the magnification factor established
by physiologic methods.
The question we were interested in, however, was not so much the density distribution of bipolar cells and photoreceptors but the size of the area in which they receive input (the dendritic field) compared to the size of their output location (axonal ramifications). Another question of interest was, whether the individual fields would overlap, and if they did, to what degree, because that would strongly influence the resolution as well as the quality of the signal transmitted to the inner retina. To use a different paradigm, a strong overlap would cause lower resolution but at the same time provide a much more stable picture owing to transmission of the same bit of information by several independent channels.
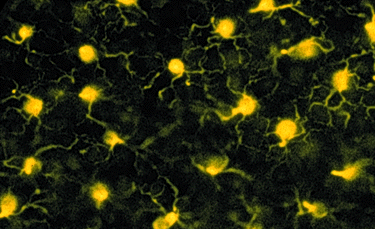
| Tangential section at the level of the outer plexiform layer which allows to examine the individual dendritic processes in detail. |
The section following the one shown above turned out to answer most
of these questions. In several sections like this, we were able to trace
and analyze in detail the processes of the bipolar cells involved in data
collection. Upon drawing a substantial number of these dendritic fields,
the unambiguous result was that each cells sends its processes out until
it reaches the center of the neighboring cells field marked by the Landolts
club. As a consequence, each photoreceptor, independent of its retinal
location, falls into the dendritic field of 3 bipolar cells belonging to
the same class.
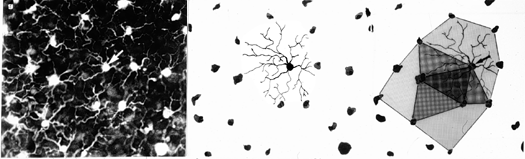
| On the left, the panel shows an original micrograph of a tangential section though the OPL, the middle panel shows the locations of the Landolt's clubs and the traced dendrites of one cell. The right panel shows the overlapping dendritic fields of three neighboring bipolar cells |
The most exciting finding at the time, however, was that there was now an extremely easy way to determine the dendritic field sizes by simply drawing the perimeter around the nearest neighbor Landolts clubs. Using this method, we could confirm that also the signal collection area of bipolar cells grows from about 350 square µm / cell in the central retinal portions to about 40.000 square µm in the periphery. Again, the same size of stimulus reported by only one cell or by more than 100 cells of the same class, depending on which part of the retina it is projected makes a huge difference in the amount of brain space that has to deal with it (assuming that each ganglion cell claims the same size territory in the brain).
.